|
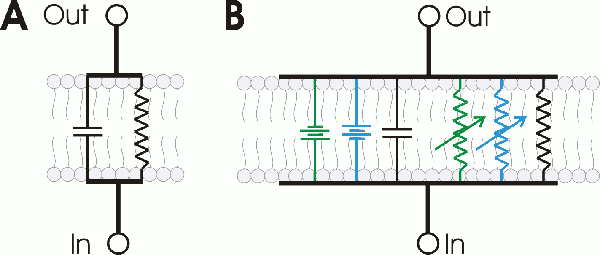
| Los potenciales de acción de membrana pueden representarse usando varios circuitos RC, cada uno representando un trozo de membrana. |
En los axones amielínicos, los potenciales de acción se propagan como una interacción pasiva entre la despolarización que se desplaza por la membrana y los canales de sodio regulados por voltaje.
Los potenciales de acción de membrana pueden representarse uniendo varios circuitos RC, cada uno representando un trozo de membrana.
Cuando una parte de la membrana celular se despolariza lo suficiente como para que se abran los canales de sodio dependientes de voltaje, los iones de sodio entran en la célula por difusión facilitada. Una vez dentro, los iones positivos de sodio impulsan los iones próximos a lo largo del axón por repulsión electrostática, y atraen los iones negativos desde la membrana adyacente.
Como resultado, una corriente positiva se desplaza a lo largo del axón, sin que ningún ion se esté desplazando muy rápido. Una vez que la membrana adyacente está suficientemente despolarizada, sus canales de sodio dependientes de voltaje se abren, realimentando el ciclo. El proceso se repite a lo largo del axón, generándose un nuevo potencial de acción en cada segmento de la membrana.
Velocidad de propagación
Los potenciales de acción se propagan más rápido en axones de mayor diámetro, si los demás parámetros se mantienen. La principal razón para que ocurra es que la resistencia axial de la luz del axón es menor cuanto mayor sea el diámetro, debido a la mayor relación entre superficie total y superficie de membrana en un corte transversal. Como la superficie de la membrana es el obstáculo principal para la propagación del potencial en axones amielínicos, el incremento de esta tasa es una forma especialmente efectiva de incrementar la velocidad de la transmisión.
Un ejemplo extremo de un animal que utiliza el aumento de diámetro de axón como regulador de la velocidad de propagación del potencial de membrana es el calamar gigante. El axón del calamar gigante controla la contracción muscular asociada con la respuesta de evasión de depredadores del animal. Este axón puede sobrepasar 1 mm de diámetro, y posiblemente sea una adaptación para permitir una activación muy rápida del mecanismo de escape. La velocidad de los impulsos nerviosos en estas fibras es una de las más rápidas de la naturaleza, para los que poseen neuronas amielínicas.
Conducción saltatoria
En axones mielínicos, la conducción saltatoria es el proceso por el que los potenciales de acción parecen saltar a lo largo del axón, siendo regenerados sólo en unos anillos no aislados (los nodos de Ranvier). La conducción saltatoria incrementa la velocidad de conducción nerviosa sin tener que incrementar significativamente el diámetro del axón.
Ha desempeñado un papel importante en la evolución de organismos más complejos cuyos sistemas nerviosos necesitan transmitir rápidamente potenciales de acción a largas distancias. Sin conducción saltatoria, la velocidad de conducción requeriría incrementos drásticos en el diámetro del axón, a tal punto que podrían resultar en la formación de sistemas nerviosos excesivamente grandes para los cuerpos que deben alojarlos.
Mecanismo detallado
El principal obstáculo para la velocidad de transmisión en axones amielínicos es la capacitancia de la membrana. La capacidad de un condensador puede disminuirse bajando el área de un corte transversal de sus placas, o incrementando la distancia entre las placas. El sistema nervioso utiliza la mielinización para reducir la capacitancia de la membrana. La mielina es una vaina protectora creada alrededor de los axones por las células de Schwann y los oligodendrocitos, células de la neuroglía que aplastan sus citoplasmas formando láminas de membrana y plasma. Estas láminas se arrollan en el axón, alejando las placas conductoras (el plasma intra y extracelular) entre sí, disminuyendo la capacitancia de la membrana.
El aislamiento resultante redunda en un conducción rápida (prácticamente instantánea) de los iones a través de las secciones mielinizadas del axón, pero impide la generación de potenciales de acción en estos segmentos. Los potenciales de acción sólo se vuelven a producir en los nodos de Ranvier desmielinizados, que se sitúan entre los segmentos mielinizados. En estos anillos hay un gran número de canales de sodio dependientes de voltaje (hasta cuatro órdenes de magnitud superior a la densidad de axones amielínicos), que permiten que los potenciales de acción se regeneren de forma eficaz en ellos.
Debido a la mielinización, los segmentos aislados del axón actúan como un cable pasivo: conducen los potenciales de acción rápidamente porque la capacitancia de la membrana es muy baja, y minimizan la degradación de los potenciales de acción porque la resistencia de la membrana es alta. Cuando esta señal que se propaga de forma pasiva alcanza un nodo de Ranvier, inicia un potencial de acción que viaja de nuevo de forma pasiva hasta que alcanza el siguiente nodo, repitiendo el ciclo.
Minimización de daños
La longitud de los segmentos mielinizados de un axón es importante para la conducción saltatoria. Deben ser tan largos como sea posible para optimizar la distancia de la conducción pasiva, pero no lo suficiente como para que la disminución en la intensidad de la señal sea tanta que no alcance el umbral de sensibilidad en el siguiente nodo de Ranvier. En realidad, los segmentos mielinizados son lo suficientemente largos para que la señal que se propaga pasivamente recorra al menos dos segmentos manteniendo una amplitud de señal suficiente como para iniciar un potencial de acción en el segundo o tercer nodo. Así se eleva el factor de seguridad de la conducción saltatoria, permitiendo que la transmisión traspase nodos en caso de que estén dañados.
Enfermedad
Algunas enfermedades afectan la conducción saltatoria y disminuyen la velocidad de desplazamiento de un potencial de acción. La más conocida de todas estas enfermedades es la esclerosis múltiple, en la que los daños en la mielina imposibilitan el movimiento coordinado.



![E_{m, K_{x}Na_{1-x}Cl } = \frac{RT}{F} \ln{ \left( \frac{ P_{Na^{+}}[Na^{+}]_\mathrm{out} + P_{K^{+}}[K^{+}]_\mathrm{out} + P_{Cl^{-}}[Cl^{-}]_\mathrm{in} }{ P_{Na^{+}}[Na^{+}]_\mathrm{in} + P_{K^{+}}[K^{+}]_{\mathrm{in}} + P_{Cl^{-}}[Cl^{-}]_\mathrm{out} } \right) }](http://upload.wikimedia.org/math/7/a/4/7a46b2fa0e3400bba919d0c1acd9b484.png)